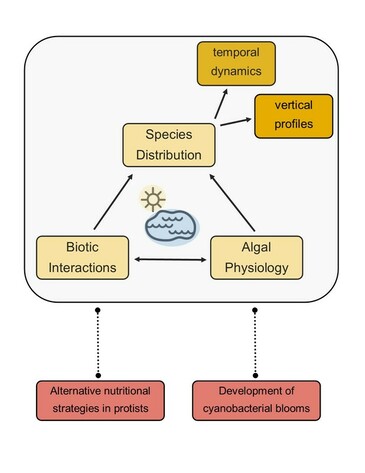
The core of my research program examines spatial and temporal distributions of planktonic protists that result from the interplay between species-specific physiology and biotic interactions. I am particularly interested in how trophic relationships between members of freshwater microbial communities (i.e. planktonic protists) contribute to the structure of communities over time, and how these patterns may change against the backdrop of anthropogenic changes in global climate and land use.
Two major themes of my research are (1) bacterivory by planktonic protists, particularly mixotrophic forms and (2) development of cyanobacterial blooms
Two major themes of my research are (1) bacterivory by planktonic protists, particularly mixotrophic forms and (2) development of cyanobacterial blooms
Physiology of Mixotrophic Protists
Protists that ingest bacteria, or bacterivores, are a keystone group in the transfer of carbon to higher trophic levels as prey for zooplankton and top-down regulators of bacteria. Bacterivory is vital to the efficiency of the microbial loop, acting as a mechanism of nutrient regeneration. Many bacterivores can function at multiple trophic levels by combining photosynthesis with ingestion of particulate matter. The utilization of mixotrophic nutrition challenges our understanding of energy flow in aquatic ecosystems. Further, mixotrophic phytoplankton function along a continuum of nutritional strategies that is often determined by species-specific physiology and environmental conditions. Inclusion of mixotrophic nutrition is lacking in mechanistic explanations of planktonic community structure, likely because it is difficult to predict the relative reliance on autotrophy and heterotrophy. It is clear that mixotrophic organisms must make physiological adjustments in the switch between trophic-modes, and such trade-offs are often dependent on the unique strategy of the mixotroph.
There is a lack of knowledge surrounding the abiotic triggers that cause mixotrophic organisms to alter their position on the mixotrophic spectrum. I have addressed this in Dinobryon, a genus of Chrysophycean algae with a near cosmopolitan distribution. Dinobryon are an effective model for facultative phagotrophy because although phototrophy represents the primary nutritional mode, they are capable of substantial rates of bacterivory (Princiotta et al 2016, Princiotta and Sanders 2017). In laboratory experiments, temperature and irradiance triggered a shift in trophic mode, where Dinobryon acquired more carbon from bacterivory under low irradiance and reduced temperature. Further experimentation revealed that this genus exhibits enhanced grazing in response to nutrient limitation.
See our publications in the Journal of Phycology here and here.
Protists that ingest bacteria, or bacterivores, are a keystone group in the transfer of carbon to higher trophic levels as prey for zooplankton and top-down regulators of bacteria. Bacterivory is vital to the efficiency of the microbial loop, acting as a mechanism of nutrient regeneration. Many bacterivores can function at multiple trophic levels by combining photosynthesis with ingestion of particulate matter. The utilization of mixotrophic nutrition challenges our understanding of energy flow in aquatic ecosystems. Further, mixotrophic phytoplankton function along a continuum of nutritional strategies that is often determined by species-specific physiology and environmental conditions. Inclusion of mixotrophic nutrition is lacking in mechanistic explanations of planktonic community structure, likely because it is difficult to predict the relative reliance on autotrophy and heterotrophy. It is clear that mixotrophic organisms must make physiological adjustments in the switch between trophic-modes, and such trade-offs are often dependent on the unique strategy of the mixotroph.
There is a lack of knowledge surrounding the abiotic triggers that cause mixotrophic organisms to alter their position on the mixotrophic spectrum. I have addressed this in Dinobryon, a genus of Chrysophycean algae with a near cosmopolitan distribution. Dinobryon are an effective model for facultative phagotrophy because although phototrophy represents the primary nutritional mode, they are capable of substantial rates of bacterivory (Princiotta et al 2016, Princiotta and Sanders 2017). In laboratory experiments, temperature and irradiance triggered a shift in trophic mode, where Dinobryon acquired more carbon from bacterivory under low irradiance and reduced temperature. Further experimentation revealed that this genus exhibits enhanced grazing in response to nutrient limitation.
See our publications in the Journal of Phycology here and here.
Mixotrophy in a Community Context: patterns with depth and season
Despite the perceived metabolic disadvantages, mixotrophs can be numerically dominant and contribute significantly to bacterivory in freshwater ecosystems. Mixotrophic protists have the potential to survive or even thrive in a variety of environmental conditions, suggesting that this is a successful nutritional strategy that allows organisms to occupy variable ecological niche space. This is particularly relevant along the vertical gradient of resources created during thermal stratification. Work in a pristine glacial-lake indicates that although seasonal patterns of mixotrophic algae were similar to those observed in other freshwater systems, abundances of trophic groups (i.e. mixotrophs, phototrophs, heterotrophs) varied within the water column (Princiotta and Sanders, 2017). Vertical mixing patterns also influenced depth-differences in grazing rates by mixotrophic nanoflagellates and their heterotrophic counterparts. Interestingly, mixotrophic nanoflagellates demonstrated increased bacterivory under ice, suggesting that mixotrophy may contribute to population maintenance over winter. However, we have also documented high abundance, yet negligable grazing, by the Chrysophyte Dinobryon during ice-cover (Princiotta et al 2023).
See our publication in Limnology and Oceanography here
See our publication in Fottea here
Despite the perceived metabolic disadvantages, mixotrophs can be numerically dominant and contribute significantly to bacterivory in freshwater ecosystems. Mixotrophic protists have the potential to survive or even thrive in a variety of environmental conditions, suggesting that this is a successful nutritional strategy that allows organisms to occupy variable ecological niche space. This is particularly relevant along the vertical gradient of resources created during thermal stratification. Work in a pristine glacial-lake indicates that although seasonal patterns of mixotrophic algae were similar to those observed in other freshwater systems, abundances of trophic groups (i.e. mixotrophs, phototrophs, heterotrophs) varied within the water column (Princiotta and Sanders, 2017). Vertical mixing patterns also influenced depth-differences in grazing rates by mixotrophic nanoflagellates and their heterotrophic counterparts. Interestingly, mixotrophic nanoflagellates demonstrated increased bacterivory under ice, suggesting that mixotrophy may contribute to population maintenance over winter. However, we have also documented high abundance, yet negligable grazing, by the Chrysophyte Dinobryon during ice-cover (Princiotta et al 2023).
See our publication in Limnology and Oceanography here
See our publication in Fottea here
Biotic Interactions Between Bacterivorous/Mixotrophic Protists and Bloom-forming Cyanobacteria: implications for toxin-production & community structure
Freshwater ecosystems face threats of deterioration through proliferation of toxin-producing cyanobacteria. Blooms of non-toxic forms also impair aquatic habitats for other organisms through reduction of photosynthetically active radiation and depletion of oxygen during decay. Multi-trophic interactions govern the formation and degradation of bloom conditions. Much attention on top-down controls of cyanobacterial blooms has been on zooplankton, yet there is a growing body of literature dedicated to the roles of planktonic protists in “grazing the bloom.” It has been suggested that phagotrophic protists can be effective grazers of cyanobacteria despite evasion through toxin production and colony formation. Work in culture demonstrates that Cryptomonas is able to ingest, and maintain growth, after co-incubation with either toxic or non-toxic Microcystis aeruginosa. Further, this work suggests that the allelopathic compounds from Microcystis may provide nutritional benefit to the mixotroph. Cryptomonas was also able to tolerate the cyanotoxin MC-LR, and concentrations of this cyanotoxin were reduced when both organisms were grown in co-culture. Our results imply that Cryptomonas has a negative impact on production or accumulation of MC-LR in Microcystis.
See our publication in a special issue of Toxins (Environmental Drivers of Algal and Cyanobacterial Toxin Dynamics) here
Freshwater ecosystems face threats of deterioration through proliferation of toxin-producing cyanobacteria. Blooms of non-toxic forms also impair aquatic habitats for other organisms through reduction of photosynthetically active radiation and depletion of oxygen during decay. Multi-trophic interactions govern the formation and degradation of bloom conditions. Much attention on top-down controls of cyanobacterial blooms has been on zooplankton, yet there is a growing body of literature dedicated to the roles of planktonic protists in “grazing the bloom.” It has been suggested that phagotrophic protists can be effective grazers of cyanobacteria despite evasion through toxin production and colony formation. Work in culture demonstrates that Cryptomonas is able to ingest, and maintain growth, after co-incubation with either toxic or non-toxic Microcystis aeruginosa. Further, this work suggests that the allelopathic compounds from Microcystis may provide nutritional benefit to the mixotroph. Cryptomonas was also able to tolerate the cyanotoxin MC-LR, and concentrations of this cyanotoxin were reduced when both organisms were grown in co-culture. Our results imply that Cryptomonas has a negative impact on production or accumulation of MC-LR in Microcystis.
See our publication in a special issue of Toxins (Environmental Drivers of Algal and Cyanobacterial Toxin Dynamics) here
